نظریهی ریاضی که دلیل انواع توهم بینایی را توضیح میدهد
توهمهایی که مخدرهای توهمزا ایجاد میکنند، مشخصاتی دارند که بین مصرفکنندهها یکی است. برای همین دانشمندان تا مدتها تصور میکردند این شباهتها به خاطر ساختار مغزی ما هستند. بعد از تقریباً یک قرن مطالعه، توضیح این دلیل شباهتها مشخصتر میشود.

در دههی ۱۹۲۰، دههها پیش از اینکه تیموتی لریTimothy Leary)) منادی و مرشدِ ضدفرهنگ در دانشگاه هاروارد به وسیلهی LSD و سایر مخدرهای توهمزا روی خودش آزمایش کند و موجهای فرهنگی از این دست به راه بیندازد، آزمایشی روی مواد مخدر توهمزا در جریان بود. یک روانشناسِ ادراکیِ جوان به نام هاینریش کلووِر (Heinrich Klüver) تصمیم گرفت سوژهی آزمایش باشد. روزی او در آزمایشگاهاش در دانشگاه مینهسوتا تکهای پیوت بلعید که در واقع نوکِ خشکشدهی کاکتوسی به اسمِ Lophophora williamsii است، و بعد با دقت چگونگی تغییر میدان بصریاش تحت تاثیر این مخدر را به ثبت رساند. توجه او به الگوهای تکرارشوندهای جلب شد که شباهتی قابلتوجه به شکلهایی داشتند که معمولا در طراحیهای غارهای کهن و در نقاشیهای خوان میرو سر و کلهشان پیدا میشد. پس نتیجه گرفت که این الگوها به احتمال قوی درونزاد [بخش لاینفک] بینایی بشر هستند. کلوور تصمیم گرفت این الگوها را در چهار دستهی متمایز به نام «ثابتهای فرمی (form constants)» طبقهبندی کند: شبکهها (از جمله تختهی شطرنج، شانهی عسل و سهگوشها)، دالانها، مارپیچها و تارعنکبوتها.
حدود پنجاه سال بعد، جک کووان (Jack Cowan) از دانشگاه شیکاگو به شیوهای ریاضیاتی شروع به بازتولید آن ثابتهای فرمی خیالانگیز کرد، با این باور که آن ثابتهای فرمی میتوانند سرنخهایی راجعبه ساختار مغزی فراهم آوردند. کووان و بارد ارمنتروت (Bard Ermentrout) دانشجوی فارغالتحصیل او در مقالهای بدیع در سال ۱۹۷۹، اینطور نوشتند که فعالیت الکتریکی نورونها در نخستین لایهی کورتکس بصری میتواند مستقیما به اشکال هندسیای ترجمه شود که آدمها به طور معمول وقتی میبینندشان که تحت تاثیر توهمزاها هستند. چندی پیش کووان در توضیح چنین اضافه کرد که «ریاضیاتی که مغز بر اساساش سیمکشی شده تنها میتواند منتج به چنین شکلهایی شود». معنی حرفش این است که آنچه در هنگام توهم میبینیم معماری شبکهی عصبی مغز را بازتاب میدهد.
اما چطور عصبکشی طبیعی کورتکسِ بینایی موجب فعالیت مغزیای میشود که به نوبهی خود باعث ایجاد توهمها میشود؟ کسی نمیداند.
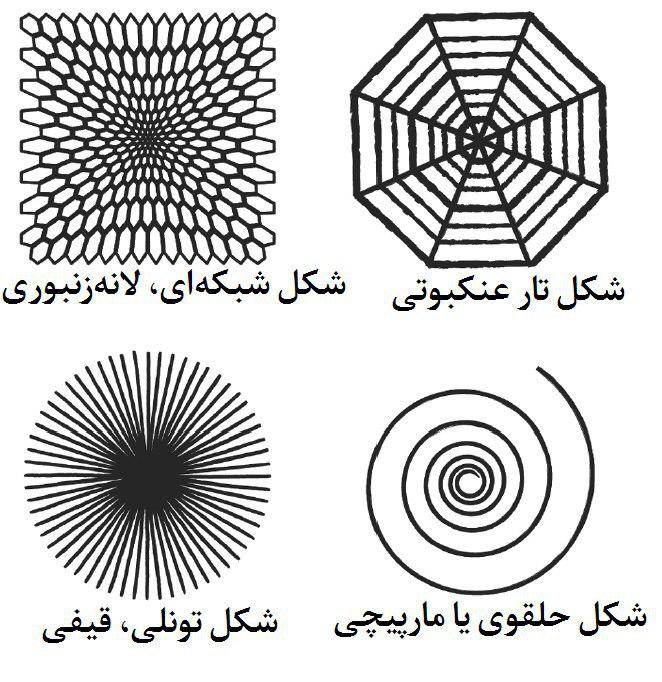
هاینریش کلوور اشکالی که تحت تاثیر مخدرهای توهمزا دید را در چهار گروه طبقهبندی کرد که به «ثابتهای فرمی» معروفاند.
اما به تازگی فرضیهای مطرح شد مبنی بر این که همه چیز زیر سر (به اصطلاح) الگوهای تورینگ است. در مقالهای به سال ۱۹۵۲، آلن تورینگِ (Alan Turing) ریاضیدان و رمزشکنِ انگلیسی مکانیسمی ریاضیاتی برای تولید بسیاری از الگوهای تکرارشونده طرح کرد که معمولا در زیستشناسی یافت میشدند ــ برای مثال بافتِ راهراهِ ببرها و گورخرها یا انواع ماهیهای خوش خطوخال یا لکههای روی بدن پلنگ. دانشمندان تا مدتها میدانستند که مکانیسم کلاسیک تورینگ نمیتواند در سیستمی رخ دهد که همچون مغز پیچیده و پر از نویز [نوفه – noise] است. اما یکی از همکاران کووان، نایجل گولدنفلدِ (Nigel Goldenfeld) فیزیکدانی از دانشگاه ایلینوی، ارباناـشمپین، جوری ایدهی تورینگ را تغییر داد که نویز را هم در نظر میگیرد. شواهدِ تجربیِ گزارششده در دومقالهی اخیر از این نظریه حمایت کردند که «تئوری تصادفی تورینگ» عامل اشکال هندسیای است که انسان در زمان توهم میبیند.
ملخهای عرقکرده
تصاویری که «میبینیم» در اصل الگوهایی هستند که به وسیلهی نورونهای تحریکشده در کورتکس بینایی ایجاد میشوند. نور از اشیایی که در میدان دیدمان قرار دارند منعکس میشود، وارد چشم میشود و در رتینا (شبکیهی چشم) متمرکز میشود؛ که پوشیده از سلولهای دریافتکنندهی نور است. این سلولها نور را به سیگنالهای الکتروشیمیایی تبدیل میکنند. این سیگنالهای الکتروشیمیایی، در شرایط عادی، به مغز میروند و نورونهای کورتکس بینایی را بر طبق الگوهایی برمیانگیزانند، این الگوها شبیه به الگوهای نوری منعکسشده از اشیایی است که در میدان دید شما قرار دارند. اما بعضی وقتها این الگوها به صورت خودانگیخته از تحریک تصادفی نورونها در کورتکس ایجاد میشوند، و بعضی مواقع یک مخدر روانگردان یا عامل تاثیرگذار دیگری عملکرد مغز نرمال را مختل میکند و تحریک تصادفی نورونها را افزایش میدهد و موجب پدیدارشدن این الگوها میشود ــــ نویزِ درونیِ پسزمینه در مقایسه با محرک بیرونی. به باور این محققان وقتی چنین اتفاقاتی در مغزمان رخ میدهد دچار توهم میشویم.
اما چرا آن شکلهایی را میبینیم که کلوور با وسواس بسیار طبقهبندی کرد؟ توضیح قابلِ قبولی که از سوی کووان، ارمنتروت و همکارانشان مطرح شده این بود که نحوهی بازنمایی میدان بینایی در نخستین ناحیهی بینایی کورتکس است که چنین الگوهایی را تولید میکند. یکی از همکاران کووان، پیتر توماس (Peter Thomas) که اکنون در دانشگاه کیس وسترن رزرو مشغول است میگوید: «اگر مغز یک نفر را بشکافید و به فعالیت سلولهای عصبی او نگاهی بیاندازید، با تصویری همانند تصویر لنز دوربین مواجه نخواهید شد». توماس توضیح میدهد که در عوض تصویر جهان همزمان که بر روی کورتکس نقش میبندد، دچار استحالهی مختصات نیز میشود. اگر فعالیت عصبی شکل راهراههای یکدرمیان نورونهای تحریکشده و تحریکنشده را به خود بگیرد، آنگاه بسته به جهتِ آن خطوط، ادراک شما هم متفاوت خواهد بود. اگر راهراهها به یک جهت متمایل باشند، حلقههایی هممرکز را میبینید. اگر راهراهها عمود به نقطهای مرکزی باشند، شما پرتوها یا شکلهای قیفمانندی را میبینید که از نقطهای مرکزی ساطع میشوند ــ همان نور معروف در انتهای تونل که عموما در تجربهی نزدیک به مرگ تجربه میکنید. و اگر راهراهها اریب باشند، الگوهای مارپیچ را میبینید.

این دیاگرام نشان میدهد که چگونه خطها در میدان بینایی (نواحی مدور در سمت چپ) به صورت خطوطی در کورتکس شیاردار ترسیم میشوند، قسمتی از کورتکس بینایی در پردازش مستقیم اطلاعات بینایی درگیر میشود.
اگر توهمهای بیناییِ هندسی مانند ثابتهای فرمی کلوور نتیجهی مستقیم فعالیت عصبی در کورتکس بینایی است پرسش اینجاست که چرا این فعالیت بهصورت خودانگیخته رخ میدهد و در این صورت چرا ما پیوسته دچار توهم نمیشویم. تئوری تصادفی تورینگ به صورت بالقوه به هردوی این پرسشها میپردازد.
آلن تورینگ در همان مقالهی کذایی اینطور بیان میکند که الگوهایی مانند لکهها نتیجهی فعلوانفعال میان دو مادهی شیمیایی است که در سرتاسر یک سیستم منتشر میشوند. این دو مادهی شیمیایی به جای اینکه مانند گاز به صورت یکنواخت در فضا منتشر شود تا چگالیاش در سرتاسر فضا یکدست شود، به میزانهای متفاوتی پخش میشوند و این امر باعث میشود نواحی متمایز با ترکیبهای شیمیایی متفاوت ایجاد کنند. یکی از این مواد شیمیایی بهعنوان فعالکننده عمل میکند که خصیصهای مشخص را بروز میدهد، مثلاً باعث بروز رنگیزهها در نقطه یا خطی مشخص میشود، در حالی که مادهی دیگر بهعنوان یک مهارکننده بیانِ مادهی شیماییِ اول، یعنی فعالکننده را مختل میکند. برای مثال دشتی از علفهای خشک و پر از ملخ را تصور کنید. اگر شما به صورت تصادفی در چند نقطه که هیچ رطوبتی ندارد آتش روشن کنید، باقی زمین نیز خواهد سوخت. اما اگر گرمای حریق باعث شود ملخهای گریزان عرق کنند، و تعریق آنها علفهای خشک اطرافشان را مرطوب کند، در این صورت شما میمانید و نقاط متناوبی از علف سالم در سرتاسر زمینی که اگر تعریق ملخها نبود کاملا میسوخت. جیمز موری، زیستشناسِ ریاضیدان این استعاره را برای توضیح مکانیسم کلاسیک تورینگ مطرح کرده است.
البته تورینگ اعتراف میکند که این مدل برای توضیح چگونگی شکلگرفتن الگوهای واقعی، بیش از حد سادهسازی شده است. برای همین هیچگاه از این مدل برای شرح یک مسئلهی زیستشناختیِ واقعی استفاده نکرد. اما این مدل چارچوبی را مطرح میکند که میتوان بر آن اتکا کرد. کووان و ارمنتورت در مقالهای در سال ۱۹۷۹ به این مسئله اشاره کردند که نورونهای مغز را نیز میتوان نوعی فعالکننده یا مهارکننده در نظر گرفت. نورونهای فعالکننده با با تحریک سولوهای سلولهای مجاور باعث تقویت پیام عصبی میشوند، در حالیکه نورونهای مهارکننده با خاموش کردن سلولهای مجاورشان پیام را خفه میکنند. این محققان متوجه شدند که نورونهای فعالکننده در کورتکس بینایی اغلب به نورونهای فعالکنندهی مجاور متصل میشدند، در حالیکه نورونهای مهارکننده تمایل دارند تا به نورونهای مهارکنندهی بسیار دورتری متصل شوند و همین امر باعث میشود شبکهی گستردهتری را شکل دهند. این یادآورِ میزان انتشارِ دو مادهی شیمیایی متفاوت در مکانیسم کلاسیک تورینگ است، و در حرف، میتواند بهصورت خودانگیخته خطوط راهراه و لکههای نورونهای فعال را پدید آورد که در سرتاسر دریای فعالیت مشتعل عصبی پراکنده شدهاند. این خطوطِ راهراه یا لکهها بسته به جهتگیریشان میتوانند ادراک شبکهای، دالانی، مارپیچی و لانهزنبوری را موجب شوند.
در حالیکه کووان تشخیص داد که کورتکس بینایی به نوعی با مکانیسم تورینگ سروکار دارد، اما مدل او توضیحی دربارهی نویز ــ یعنی تحریک ناگهانی و انفجاریِ نورونها ــ نداشت، نویزی که به نظر میرسید مانع شکلگیری الگوهای تورینگ میشود. در همین حین، گولدنفلد و دیگر محققان ایدههای تورینگ را در بومشناسی، در رابطه با دینامیک شکارـشکارچی به کار گرفتند. در آن سناریو، شکارها همچون فعالکنندهها عمل میکنند و به دنبال تولید مثل و افزایش تعدادشان هستند، در حالیکه شکارچیها همچون مهارکنندهها با شکارکردن جمعیت شکار را کنترل میکنند. بنابراین، آنها با یکدیگر الگوهای فضایی تورینگی را شکل میدهند. گلدنفلد در حال بررسی تأثیر نوسانات تصادفی جمعیتهای شکار و شکارچی بر روی این الگوها بود. او از مطالعات کووان در عصبشناسی خبر داشت و کمی بعد متوجه شد که دانش او میتواند در آن زمینه به کار گرفته شود.
خانههایی با چشم و آرواره
گولدنفیلد (Goldenfeld)، فیزیکدان کارآموخته در زمینهی مادهْ چگال (condensed matter) مجذوب پژوهشهای بینارشتهای میشود و تکنیکها و مفاهیمِ فیزیک و ریاضی را در زیستشناسی و بومشناسی تکاملی به کار میگیرد. تقریبا ده سال پیش، او به همراه تام باتلر (Tom Butler) دانشجوی فارغالتحصیلاش در آن دوره به این مسئله فکر میکردند که نوسانات تصادفی محلی در جمعیت شکار و شکارچی چگونه توزیع فضاییشان را دگرگون میکند، برای نمونه اگر گلهای از گوسفندان مورد حملهی گرگ قرار بگیرند. گلدنفیلد و باتلر دریافتند که وقتی جمعیت گوسفندان نسبتا کم است، نوسانات تصادفی میتوانند تأثیرات به سزایی داشته باشند، حتا تا جایی که موجب انقراض کل جمعیت شوند. معلوم شد که مدلهای بومشناختی نه صرفاً رفتار میانگین جمعیتها بلکه باید نوسانات تصادفی را در نظر بگیرند. گلدنفیلد میگوید «از قبل میدانستم که چطور محاسبات نوسانی را برای شکلگیری الگوها به کار ببرم. پس مشخصاً قدم بعدی به کار بردن محاسبات نوسانی برای مسئلهی توهم بود.»
در مغز، به جای جمعیتهای گوسفند و گرگ جمعیتی از نرونها هستند که با نوسانات تصادفی روشن و خاموش میشوند. اگر یک نرون فعالکننده تصادفاً روشن شود، میتواند نرونهای مجاور را نیز روشن کند. برعکس، هنگامی که یک نرون مهارکننده، تصادفی روشن شود، منجر به خاموششدن نرونهای مجاور میشود. با توجه به اینکه اتصالات نرونهای مهارکننده با نرونهایی است که در فواصل دورتر قرار دارند، اگر هر یک از این سیگنالها تصادفاً فعال شوند سریعتر از سیگنالهای تهییجی تصادفی منتشر میشوند ــ این دقیقاً همان چیزی است که یک مکانیسم تورینگی لازم دارد. مدلهای گلدنفلد به این مسئله اشاره دارد که راهراههای نرونهای فعال و غیرفعال در یک الگوی تورینگی شکل خواهند گرفت. او اینها را «الگوهای تصادفی تورینگ» نامید.
هر چند کورتکس بینایی برای اینکه درست عمل کند باید از سوی یک محرک خارجی برانگیخته شود و نه به وسیلهی نوسانات نویزی داخلی خودش. چه چیزی از شکلگرفتن دائمی الگوهای تصادفی تورینگ که موجب توهم مداوم میشوند جلوگیری میکند؟ گلدنفلد و همکاراناش اینطور استدلال میکنند که هر چند تحریک شدن نورونها میتواند تصادفی باشد، اما اتصالات بین آنها تصادفی نیستند. هرچند اتصالات کوتاهبرد بین نورونهای تهییجی متداول و معمول است، اما اتصالات دوربرد بین نورونهای مهارکننده نامتراکم و پراکنده است، و گلدنفلد گمان میکند که این مسئله به خنثاکردنِ انتشار سیگنالهای تصادفی کمک میکند. او و همکاراناش این فرضیه را با ساخت دو مدل شبکهی عصبیِ مجزا آزمایش کردند. یکی از مدلها بر اساس سیمکشی واقعی کورتکس بینایی، و مدلِ دیگر شبکهای عام با اتصالات تصادفی بود. در مدل عام، عملکرد بینایی نرمال عملاً از بین رفت زیرا تحریک تصادفی نورونها در خدمت تقویت اثر تورینگ برآمد. گلدنفلد میگوید: «یک کورتکس بینایی که بهصورت تصادفی سیمپیچی شده به ضرب هجوم توهمها آلوده و خراب میشود». هر چند، در مدل واقعگرایانه از کورتکس، نویز درونی بهصورت موثری فرونشانده میشد.

نایجل گلدنفلد، فیزیکدانی در دانشگاه ایلینوی، ارباناـشمپیون، این فرضیه را طرح میکند که مکانیزم تورینگ مبنای توهمهای بینایی است.
گلدنفلد خاطرنشان میشود که طی تکامل یک ساختار شبکهای خاص انتخاب شده و بالا آمده. این شبکه از بروز الگوهای توهمی جلوگیری میکند: پراکندگیِ اتصالات مابین نورونهای مهارکننده، مانع از حرکت سیگنالهای مهارکننده در مسافتهای طولانی میشود، و تئوری تصادفی تورینگ، ادراکِ قیفی، لانهزنبوری، مارپیچی و الخ را مختل میکند. بنابراین غالب الگوهایی که روی شبکه پخش میشوند، از محرک خارجی نشأت میگیرند ــ که این برای بقاء چیز خوبی است، زیرا شما باید یک مار تشخیص دهید، نه اینکه شکل تقریباً مارپیچ مار حواستان را پرت و مختل کند.
توماس میگوید: «اگر سرتاسرِ کورتکس از این اتصالات مهارکنندهی دوربرد ساخته شده بود، آنگاه کورتکس بیشتر تمایل داشت به جای پردازش ورودی بینایی الگوهایی توهمزا تولید کند. در این صورت ما با یک فاجعه مواجه میشدیم و ممکن نبود زنده بمانیم». زیرا اتصالات مهارکنندهی دوربرد پراکنده هستند، «این مدلها الگوهای خودانگیخته تولید نمیکنند مگر اینکه شما با القای اثر مخدرهای توهمزا آنها را مجبور کنید.»
آزمایشها نشان دادهاند که توهمزاهایی مانند LSD ظاهراً مکانیسمهای نرمالِ پالایش و تصفیه را که مغز به کار میبرد مختل میکنند. احتمالاً چنین توهمزاهایی اتصالات مهارکنندهی دوربرد را افزایش میدهند و بنابراین به سیگنالها مجال میدهند تا به صورت تصادفی به واسطهی «اثرِ تصادفی تورینگ» تقویت شوند.
گلدنفلد و همکاراناش هنوز نظریهشان دربارهی توهمهای بینایی را بهشیوهای تجربی آزمایش نکردهاند، اما طی سالهای اخیر شواهد انکارناپذیری به دست آمده مبنی بر اینکه الگوهای تصادفی تورینگ قطعاً در سیستمهای زیستشناختی رخ میدهند. در حدود سالِ ۲۰۱۰، گلدنفلد از پژوهشهای رونالد وایس (Ronald Weiss) مطلع شد. وایس یک زیستشناس ترکیبی در موسسهی تکنولوژی ماساچوست است و سالها کوشیده است تا چهارچوب نظری مناسبی برای توضیح برخی نتایجِ آزمایشگاهی و تجربیِ جالب توجه پیدا کند.
سالها پیش، وایس و تیماش بیوفیلم [ماتریکسی از میکروارگانیسمها که یک سطح را تشکیل میدهند] باکتریایی که از لحاظ ژنتیکی مهندسی شده بودند را پرورش دادند تا به وسیلهی آنها یکی از دو ملکول سیگنالدهندهی متفاوت را تولید کنند. در این آزمایش که به منظور نشاندادن رشد الگوهای کلاسیک تورینگ انجام شد، آنها ملکولهای سیگنالدهنده را با نشانهای فلورسنت مشخص کردند، طوریکه فعالکنندهها با نور قرمز و مهارکنندهها با نور سبز میدرخشیدند. گرچه این آزمایش با یک بیوفیلم یکدست آغاز شد، اما بعد از مدتی الگویی تورینگی پدیدار شد آن هم به صورت خالخالهای قرمز که در سرتاسر یک ردیفِ سبز پراکنده شده بودند. هر چند این نقاط قرمز در مقایسه با (مثلاً) خالهای یک پلنگ بهنحوی بسیار تصادفیتر مستقر شده بودند. آزمایشهای بیشتر در این زمینه نیز شکست خوردند.
هنگامی که گلدنفلد دربارهی این آزمایشها شنید، احتمال داد که چه بسا دادههای وایس میتوانست از دیدگاه تصادفی نیز دیده شود. وایس گفت «به جای اینکه تلاش کنیم تا الگوها هر چه منظمتر و کم نویزتر شوند، از خلال همکاریمان با نایجل متوجه شدیم که این الگوها در واقع الگوهای تصادفی تورینگ هستند.» سرانجام در ماه گذشته، وایس، گلدنفلد و سایر همکارانشان، بعد از گذشت هفده سال از شروع این پژوهش، مقالههایشان را در مجلهی علمی (PNAS (Proceedings of the National Academy of Sciences منتشر کردند.
بیوفیلمها به الگوهای تصادفی تورینگ سر و شکل دادند زیرا بیان ژن یک فرآیند نویزی است. بر طبق نظر جوئل استاوانس (Joel Stavans) در موسسهی علمی وایزمن در اسرائیل، نویز مذکور موجب ایجاد تمایز در میان سلولهاست، سلولهایی که با وجود اطلاعات ژنتیکی که دارند میتوانند بهشیوهای متفاوت رفتار کنند. استاوانس و همکاراناش در کتابی که به تازگی منتشر کردهاند، بررسی کردهاند که نویز چگونه در بیان ژن میتواند به الگوهای اتفاقی تورینگ در سیانوباکتری ــ ارگانیسمهای باستانی که حجم عظیمی از اکسیژن برای کرهی زمین تولید کردهاند ــ منجر شود. محققان، آنابنا، نوعی از سیانوباکتریا را مورد مطالعه قرار دادند که ساختار سلولی سادهای دارند، این آنابناها در رشتهای طویل به یکدیگر متصل شدهاند. سلولهای آنابنا میتوانند یکی از این دو فعالیت را به صورت تخصصی انجام دهند: فتوسنتز، یا تبدیل نیتروژن جو زمین به پروتئینها. برای نمونه، یک آنابنا در آنچه ظاهراً الگوی تصادفی تورینگ است، احتمالاً دارای یک سلول تثبیتکنندهی نیتروژن، و سپس ده تا پانزده سلول فتوسنتز، و بعد از آن یک سلول تثبیتکنندهی نیتروژن دیگر و همینطور الخ است. در این مورد، فعالکننده یک پروتئین است که حلقهی بازخورد مثبتی را برای تولید پروتئینهای بیشتری از این دست ایجاد میکند. در عین حال، این پروتئین میتواند پروتئینهایی از نوع دیگر تولید کند که به سلولهای همسایه نشت میکنند و جلوی تولید اولین پروتئین را میگیرند. این خصیصهیِ اصلی مکانیسم تورینگ است: یک فعالکننده و یک مهارکننده با هم به رقابت میپردازند. در آنابنا، نویز این رقابت را جلو میبرد.
به گفتهی محققان این که «فرآیندهای تصادفی تورینگ» در این دو روند زیستشناختی در کارند، احتمال این که روند مشابهی در کورتکس بینایی هم در کار باشد را بالاتر میبرد. یافتهها همچنین نشان میدهند که نویز چگونه نقشی محوری در ارگانیسمهای زیستشناختی بازی میکند. وایس میگوید: «رابطهی مستقیمی میان نحوهی برنامهریزی یک کامپیوتر به وسیلهی ما» و نحوهی عملکردِ سیستمهای زیستشناختی وجود ندارد. «زیستشناسی نیازمند چارچوبها و اصول طراحی متفاوتی است که نویز یکی از آنهاست.»
برای درک سازوکار توهم هنوز راه درازی در پیش است. ژان پل سارتر در ۱۹۳۵ در پاریس مسکالین را تجربه کرد و متوجه شد که مصرف آن ادراک بیناییاش را برای هفتهها معوج کرده است. در تجربهی او، خانهها «سطوحی چشمچران به نظر میرسیدند، سراسر مملو از چشمها و آروارهها»، سیمای ساعتها به جغدها میمانستند، و او خرچنگهایی را دید که در تمام مدت به دنبال او هستند. توهمهای سارتر در نسبت با ثابتهای فرمیِ سادهی کلوور توهمهای بسیار سطح بالاتری هستند. ارمنتروت میگوید «مراحل اولیهی توهم بینایی که همان مشاهدهی الگوهای هندسی است، بسیار ساده هستند». اما وقتی عملکردهای شناختیِ سطح بالاتری مثل حافظه درگیر میشوند، «شما شروع به دیدن توهمهای پیچیدهتری میکنید و میکوشید آنها را بفهمید. فکر میکنم، در اینگونه توهمها بیشتر نواحی اصلیتر مغز برانگیخته میشوند و هر آنچه میبینید بروزِ خودانگیختهی [حافظههای ذخیرهشده] است.»
در دههی ۲۰ کلوور همچنین با سوژههایی کار میکرد که توهمهای لامسهای را گزارش داده بودند، توهمهایی مثلِ خزیدنِ تارعنکبوتها روی پوست. ارمنتروت فکر میکند که این توهمهای لامسهای با یک ثابتِ فرمیِ تارعنکبوتی جور درمیآید، ثابتفرمی تارعنکبوتی که بر کورتکس سوماتوسنسوری (حسهای تنانه) منطبق میشود. همین فرآیندها در رابطه با کورتکس شنوایی نیز رخ میدهند و میتوانند نه تنها شرحی بر توهمهای شنوایی بلکه توضیحی دربارهی پدیدههایی مانند زنگزدن گوش نیز باشند. با توجه به اینکه سیمکشی مشابهی در سرتاسر مغز وجود دارد، پس اگر نظریهی توهمها «در رابطه با بینایی کار کند، در موردِ همهی حواس دیگر نیز کار خواهد کرد.»

پیشنهاد کتاب
-

یفرن دوم
-
گریخته: هفتروایت در باب مرگ
-
ماورا: سلسله جنایتهای بین کهکشانی
-
شومنامهی تبر نقرهای




آخرین دیدگاهها
نظر خود را بنویسید:
پیشنهاد کتاب
-
یفرن دوم
-
گریخته: هفتروایت در باب مرگ
-
ماورا: سلسله جنایتهای بین کهکشانی
-
شومنامهی تبر نقرهای